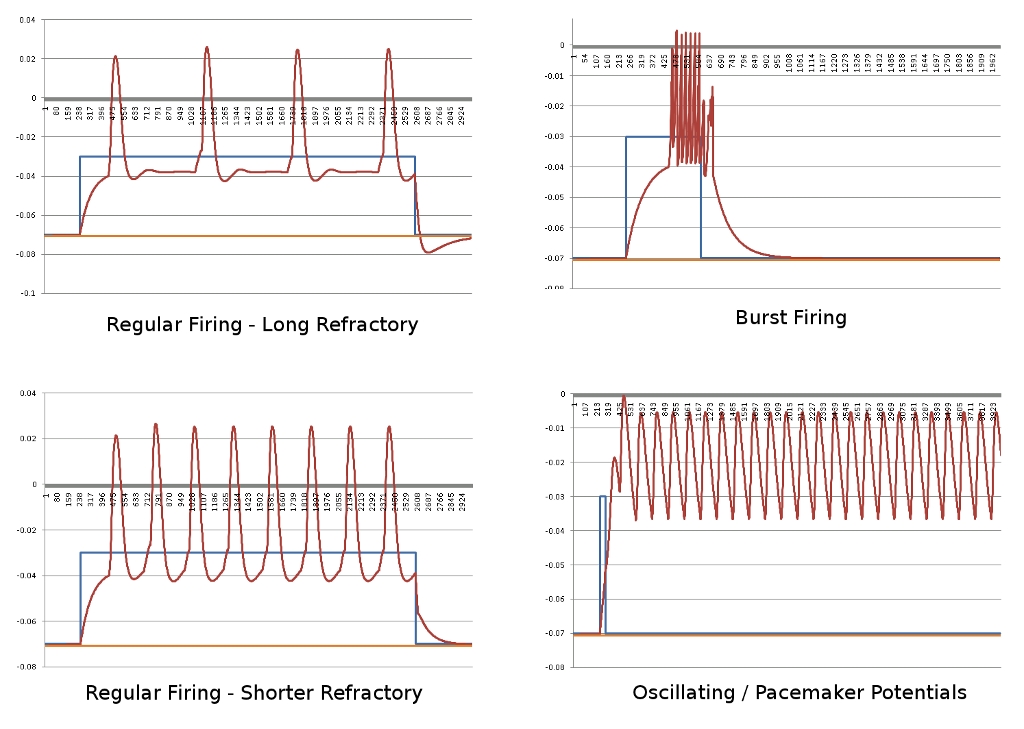
Progress marches forward on the Neural Emulator front. I’ve currently fleshed out the functionality as described by the cable equation, that describes how voltage/current flows down neural structures. This will allow adjacent sections of the cellular membrane to propagate changes in potential, thereby properly emulating the action potential. Before I can advance at all, I need to ensure that the action potential sequence models properly, since this is such core functionality.
Voltage Propagation
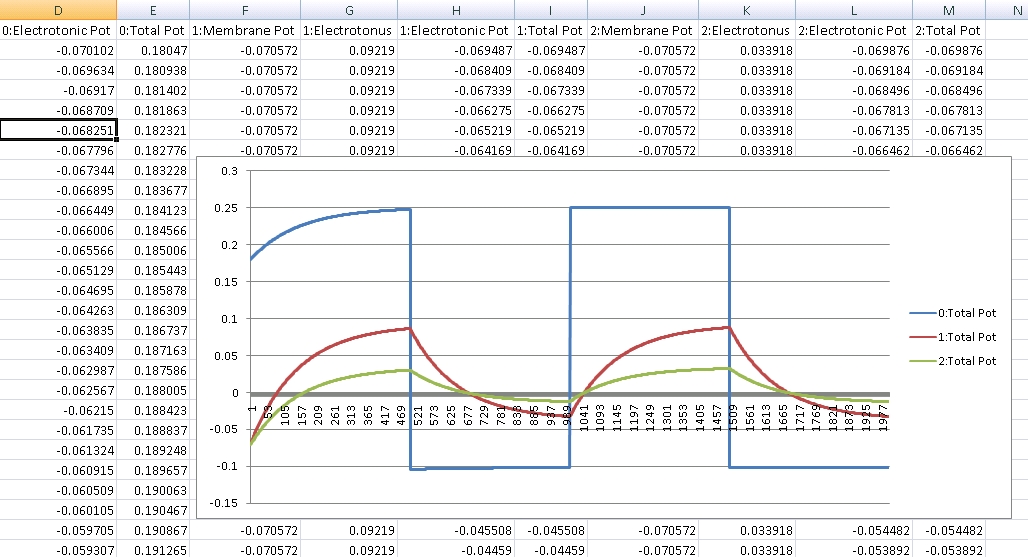
In the following graph, I’ve setup a neuron consisting of 4 structures. For the purposes of this test, it doesn’t really matter what the structures themselves are, but you could think of it as 4 sections of a fiber in a dendritic arbor. They all start out with the same intra and extracellular ionic concentrations, membrane permeability, and size. They are arranged linearly, where structure 0 is connected to 1, which is connected to 2, which is connected to 3. In this experiment, I increased in the extracellular concentration of Sodium surrounding structure 0. The graph shows both the local potential (potential for the cell membrane when isolated from adjacent membranes), as well as the total potential (when accounting for adjacent membranes).
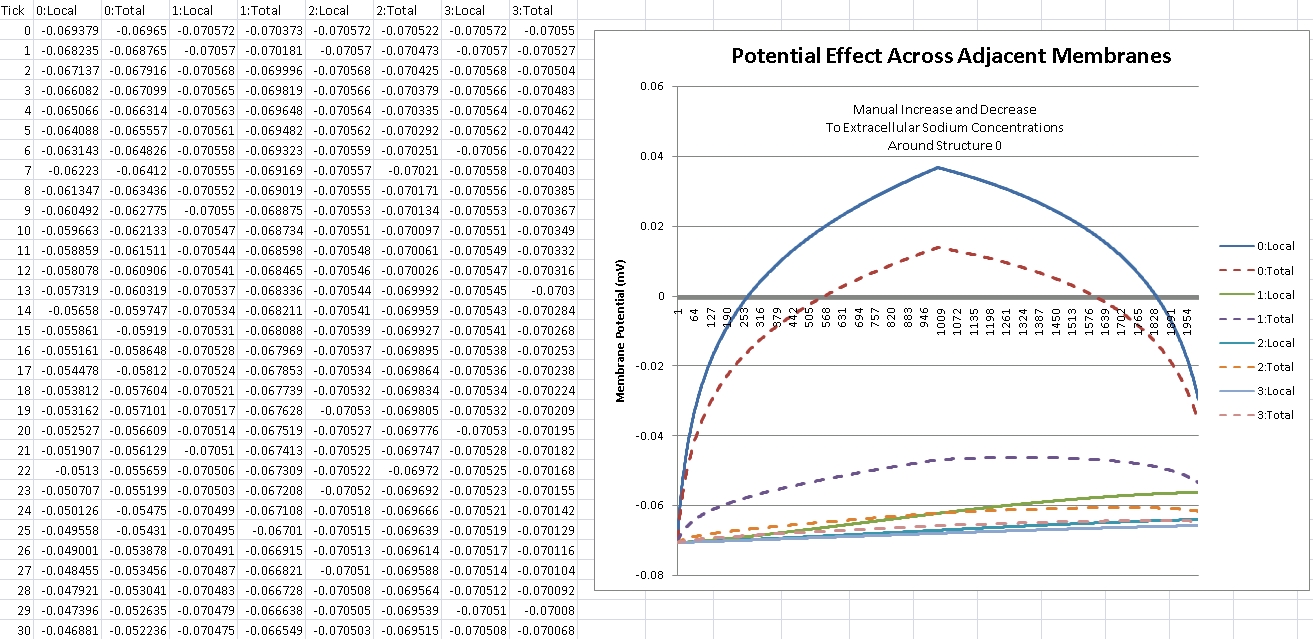
As can be seen, as we increase the extracellular Sodium concentration, the cell membrane of structure 0 depolarizes as the local potential goes positive. Though the Sodium concentration surrounding the adjacent structures has (mostly) not changed, as can be seen by their local potentials, their total potential increases accordingly due to their proximity to structure 0. The closer they are (structure 1 is the closest), the more their membrane potential is affected. The effects of such are calculated by voltage difference, connecting membrane area, and distance between them. So this test came out successful.
Hodgekin-Huxley
In addition, I’m about half way finished with integrating the Hodgekin-Huxley model and associated equations in with calculating the permeability of gated ion channels, specifically for voltage and inactivation gates. This will ensure that the ion permeability adjusts correctly depending on the membrane potential. However, before I was able to move forward on HH, I needed to ensure membrane potentials were propagating properly, which is why the work above was important. More on this soon!